Вам вже відомі такі поняття, як «середовище» і «середовище життя». Потрібно навчитися їх розрізняти. Що ж таке середовище життя?
Середовище життя - частина природи з особливим комплексом факторів, для існування в якій у систематичних груп організмів сформувалися подібні адаптації.
На Землі можна виділити чотири основні середовища життя: водне, наземно-повітряне, ґрунтове, живий організм.
Водне середовище
Водне середовищежиття характеризується високою щільністю, особливими температурним, світловим, газовим та сольовим режимами. Організми, що мешкають у водному середовищі, називаються гідробіонтами(Від грец. hydor- Вода, bios- Життя).
Температурний режим водного середовища
У воді температура змінюється меншою мірою, ніж на суші, через високу питому теплоємність і теплопровідність води. Підвищення температури повітря на 10 °C спричиняє підвищення температури води на 1 °C. З глибиною температура поступово знижується. На більших глибинах температурний режим відносно постійний (не вище за +4 °C). У верхніх шарах спостерігаються добові та сезонні коливання (від 0 до +36 °C). Оскільки у водному середовищі температура змінюється у вузькому діапазоні, то для більшості гідробіонтів потрібна стабільна температура. Для них згубні навіть невеликі відхилення температури, спричинені, наприклад, скиданням підприємств теплих стічних вод. Гідробіонти, здатні існувати при великих коливаннях температури, зустрічаються лише у дрібних водоймах. Через невеликий об'єм води в цих водоймах спостерігаються значні добові та сезонні перепади температури.
Світловий режим водного середовища
Світла у воді менше, ніж у повітрі. Частина сонячних променіввідбивається від її поверхні, а частина поглинається товщою водою.
День під водою коротший, ніж на суші. Влітку на глибині 30 м він становить 5 год, а на глибині 40 м – 15 хв. Швидке зменшення світла з глибиною пов'язане з його поглинанням водою.
Кордон зони фотосинтезу в морях знаходиться на глибині близько 200 м. У річках вона коливається від 1,0 до 1,5 м і залежить від прозорості води. Прозорість води в річках та озерах сильно знижується через забруднення зваженими частинками. На глибині понад 1500 м світло практично відсутнє.
Газовий режим водного середовища
У водному середовищі вміст кисню в 20-30 разів менше, ніж у повітрі, тому він є лімітуючим фактором. Кисень надходить у воду за рахунок фотосинтезу водних рослин та здатності кисню повітря розчинятися у воді. При перемішуванні води вміст кисню у ній зростає. Верхні шари води багатші киснем, ніж нижні. За дефіциту кисню спостерігаються замори (масова загибель водних організмів). Зимові замори бувають, коли водоймища покриваються льодом. Літні - коли через високої температуриводи зменшується розчинність кисню. Причиною може бути підвищення концентрації токсичних газів (метану, сірководню), що утворюються при розкладанні відмерлих організмів без доступу кисню. Через мінливість концентрації кисню більшість водних організмів щодо нього є еврібіонтами. Але є й стенобіонти (форель, планарію, личинки поденок і потічків), які не переносять нестачі кисню. Вони є індикаторами чистоти води. Вуглекислий газ розчиняється у воді в 35 разів краще кисню, і його концентрація в ній у 700 разів вища, ніж у повітрі. У воді CO2 накопичується завдяки дихання водних організмів, розкладу органічних залишків. Вуглекислий газ забезпечує фотосинтез та використовується при утворенні вапняних скелетів безхребетних.
Сольовий режим водного середовища
Солоність води відіграє важливу роль у житті гідробіонтів. Природні води за вмістом солей поділяють на групи, подані в таблиці:
У Світовому океані солоність становить середньому 35 г/л. Найвищий вміст солей у солоних озерах (до 370 г/л). Типові жителі прісних та солоних вод є стенобіонтами. Вони не переносять коливань солоності води. Еврібіонтів порівняно небагато (лящ, судак, щука, вугор, колюшка, лосось та ін.). Вони можуть жити як у прісній, так і в солоній воді.
Адаптації рослин до життя у воді
Всі рослини водного середовища називаються гідрофітами(Від грец. hydor- Вода, phyton- Рослина). У солоних водах мешкають лише водорості. Тіло у них не поділено на тканини та органи. До зміни складу сонячного спектру залежно від глибини водорості пристосувалися шляхом зміни складу пігментів. При переході від верхніх шарів води до глибинних забарвлень водоростей змінюється в послідовності: зелені — бурі — червоні (глибоководні водорості).
Зелені водорості містять зелений, помаранчевий та жовтий пігменти. Вони здатні до фотосинтезу за досить високої інтенсивності сонячного світла. Тому живуть зелені водорості в дрібних прісних водоймищах або на морському мілководді. До них відносяться: спірогіра, улотрикс, ульва та ін У бурих водоростей, крім зеленого, містяться бурий і жовтий пігменти. Вони здатні вловлювати менш інтенсивне сонячне випромінюванняна глибині 40-100 м. Представниками бурих водоростей є фукус і ламінарія, що мешкають лише морях. Червоні водорості (порфіра, філофора) можуть жити на глибині понад 200 м. Крім зеленого, вони мають червоний та синій пігменти, здатні вловлювати навіть незначне світло на великій глибині.
У прісних водоймах у стеблах вищих рослинслабо розвинена механічна тканина. Наприклад, якщо витягти з води латаття біле або кубочок жовте, то їх стебла поникають і не здатні підтримувати квітки у вертикальному положенні. Опорою для них є вода за рахунок її високої щільності. Адаптацією до нестачі кисню у воді є наявність в органах рослин аеренхіми (повітряної тканини). Мінеральні речовини знаходяться у воді, тому слабо розвинені провідна та коренева системи. Коріння може взагалі бути відсутнім (ряска, елодея, рдест) або служити для закріплення в субстраті (рогоз, стрілолист, шапка). Кореневих волосків на коренях немає. Листя частіше тонкі і довгі або сильно розсічені. Мезофіл не диференційований. Продихання у плаваючого листя знаходиться на верхній стороні, а у занурених у воду - відсутні. Для деяких рослин характерна наявність листя різної форми (гетерофілія) залежно від того, де вони знаходяться. У латаття і стрілоліста форма листя у воді та на повітрі різна.
Пилок, плоди та насіння водних рослин пристосовані до поширення водою. Вони мають пробкові вирости або міцні оболонки, що запобігають попаданню води всередину та загниванню.
Адаптації тварин до життя у воді
У водному середовищі тваринний світ багатший, ніж рослинний. Завдяки незалежності від сонячного світла тварини заселили всю товщу води. На кшталт морфологічних і поведінкових адаптацій їх поділяють такі екологічні групи: планктон, нектон, бентос.
Планктон(Від грец. planktos- ширяючий, блукаючий) - організми, що живуть в товщі води і пересуваються під дією її струму. Це дрібні ракоподібні, кишковопорожнинні, личинки деяких безхребетних. Усі їх адаптації спрямовані на підвищення плавучості тіла:
- збільшення поверхні тіла за рахунок сплющування та подовження форми, розвитку виростів та щетинок;
- зменшення щільності тіла у зв'язку з редукцією кістяка, наявністю жирових крапель, бульбашок повітря, слизових чохлів.
Нектон(Від грец. nektos- Плавучий) - організми, що живуть в товщі води і ведуть активний спосіб життя. Представниками нектону є риби, китоподібні, ластоногі, головоногі молюски. Протистояти течії їм допомагають адаптації до активного плавання та зменшення тертя тіла. Активне плавання досягається з допомогою добре розвиненої мускулатури. При цьому можуть використовуватися енергія струменя води, що викидається, згинання тіла, плавники, ласти і т. д. Зменшенню тертя тіла сприяють адаптації: обтічна форма тіла, еластичність шкірних покривів, наявність на
шкірі луски та слизу.
Бентос(Від грец. benthos- Глибина) - організми, що мешкають на дні водойми або в товщі донного грунту.
Адаптації бентосних організмів спрямовані на зменшення плавучості:
- обтяження тіла за рахунок раковин (молюски), хітинізованих покривів (раки, краби, омари, лангусти);
- закріплення на дні за допомогою органів фіксації (присоски у п'явок, гачки у личинок потічка) або сплощеного тіла (скати, камбала). Деякі представники зариваються в ґрунт (багатощетинкові черв'яки).
В озерах та ставках виділяють ще одну екологічну групуорганізмів – нейстон. Нейстон- організми, пов'язані з поверхневою плівкою води і мешкають постійно або тимчасово на цій плівці або до 5 см у глибину від її поверхні. Їхнє тіло не змочується, оскільки його щільність менша за щільність води. Особливо влаштовані кінцівки дозволяють пересуватися поверхнею води, не занурюючись (клопи водомірки, жуки вертячки). Своєрідною групою водних організмів є також перифітон- організми, що утворюють на підводних об'єктах плівку обростання. Представниками перифітону є водорості, бактерії, протисти, ракоподібні, двостулкові молюски, малощетинкові черв'яки, мошанки, губки.
На планеті Земля виділяють чотири основні середовища життя: водне, наземно-повітряне, грунтове і живий організм. У водному середовищі лімітуючим фактором є кисень. За характером адаптацій водних жителів поділяють на екологічні групи: планктон, нектон, бентос.
Водні рослинимають суттєві відмінності від наземних рослинних організмів. Так, здатність водних рослин поглинати вологу та мінеральні солі безпосередньо з навколишнього середовища відбивається на їх морфологічній та фізіологічній організації. Характерним для водних рослин є слабкий розвиток провідної тканини та кореневої системи. Коренева система є головним чином для прикріплення до підводного субстрату і не виконує функції мінерального живлення та водопостачання, як у наземних рослин. Живлення водних рослин здійснюється всією поверхнею їх тіла. Значна щільність води дає можливість проживання рослин у всій її товщі. У нижчих рослин, що заселяють різні шари і ведуть плаваючий спосіб життя, для цього є спеціальні придатки, які збільшують їх плавучість і дозволяють утримуватися у зваженому стані. Вищі гідрофіти мають слабко розвинену механічну тканину. У їхньому листі, стеблах, коренях розташовуються повітроносні міжклітинні порожнини, що збільшують легкість і плавучість зважених у воді та плаваючих на поверхні органів, що також сприяє омиванню внутрішніх клітин водою з розчиненими в ній солями та газами. Гідрофіти відрізняються великою поверхнею листя при малому загальному обсязі рослини, що забезпечує їм інтенсивний газообмін при нестачі розчиненого у воді кисню та інших газів.
У ряду водних організмів розвинена різнолиста, або гетерофілія. Так, у сальвінії занурене листя забезпечує мінеральне харчування, а плаваючі - органічне.
Важливою особливістю адаптації рослин до проживання у водному середовищі є і те, що листя, занурене у воду, як правило, дуже тонке. Часто хлорофіл у них розташовується в клітинах епідермісу, що сприяє посиленню інтенсивності фотосинтезу при слабкому висвітленні. Такі анатомо-морфологічні особливості найбільш чітко виражені у водних мохів, валіснерії, рдестів.
Від вимивання у водних рослин із клітин мінеральних солей або вилуговування захистом є виділення спеціальними клітинами слизу та утворення ендодерми з більш товстостінних клітин у вигляді кільця.
Відносно низька температура водного середовища обумовлює відмирання вегетуючих частин у занурених у воду рослин після утворення зимових бруньок і заміну літніх тонких нижніх листків жорсткішими і короткими зимовими. Низька температураводи негативно позначається на генеративні органиводних рослин, а висока її щільність ускладнює перенесення пилку. У зв'язку з цим водні рослини інтенсивно розмножуються вегетативним шляхом. Більшість плаваючих на поверхні та занурених рослин виносять квітконосні стебла у повітряне середовище та розмножуються статевим шляхом. Пилок розноситься вітром та поверхневими течіями. Плоди та насіння, які утворюються, також поширюються поверхневими течіями. Це явище зветься гідрохорії. До гідрохорних відносяться не тільки водні, а також багато прибережні рослини. Їхні плоди мають високу плавучість, тривалий час перебувають у воді і не втрачають при цьому схожість. Наприклад, водою переносяться плоди та насіння стрілоліста, сусака, частухи. Плоди багатьох осок поміщені у своєрідні мішечки з повітрям і розносяться водними течіями.
Організм постійно піддається дії мінливих чинників довкілля. Було вже зазначено, що це призводить до формування екологічної валентності виду, що є відображенням ступеня впливу екологічних факторів. Отже, аналіз угруповань організмів здатний давати об'єктивну інформацію про стан навколишнього середовища. Ми розглянемо здатність організмів відбивати параметри різних середовищ.
1. загальні положення. Відомо, що сильніше змінюється цей параметр середовища у просторі чи часі, тим більше екологічне значення він має для організмів. Істотну роль при цьому відіграють і ступінь регулярності впливу даного фактора на організми, регулярність фактору змінності в часі. Відповідно до цього серед мінливих факторів можна розрізняти ті, що змінюються закономірно періодично і без закономірної періодичності. Фактори, що змінюються періодично, зумовлюють формування у видів характерних їм біологічних циклів (добових, сезонних, річних тощо.), специфічних типів динаміки чисельності деяких інших видів ознак. Чинники, змінюються без закономірної періодичності, переважно впливають поширення виду та її чисельність. Чинники, що мають періодичний і регулярний характер, можна поділити на первинні та вторинні. До перших відносяться дії, які є прямим наслідком ритміки геофізичних циклів. Вторинні - це впливи, що є непрямим наслідком ритміки геофізичних циклів (наприклад, паводкові та міжпаводкові періоди).
Особини кожного виду характеризуються певним типомобміну речовин та енергії, без збереження якого не можуть успішно розвиватися. Якщо стан середовища такий, що організму загрожує порушення балансу обміну речовин та енергії, він або змінює становище у просторі, наводячи себе наскільки можна оптимальні умови середовища, або змінює інтенсивність обміну і навіть характер останнього.
2. Оптимізація екологічних умов. Оптимальними називаються умови, у яких організм із найменшими енергетичними витратами зберігає характерний йому тип обміну речовин. Оптимальні значення будь-якого чинника стосовно різних функцій організму різні, тому оптимум щодо організму загалом – поняття інтегральне. Оптимальні умови – це не ті, що забезпечують найкращі умовидля протікання окремих функцій, а ті, при яких сумарний ефект протікання всіх функцій є найкращим з метою забезпечення біологічного процвітання виду. Оптимальне дозування чинника кожному за організму є величину не постійну, а змінну, залежить від стану самого організму і від усієї суми умов, у яких живе.
Не всі фактори водного середовища відіграють рівну роль у житті організмів. У зв'язку з цим вони поділяються на першорядні та другорядні. До найважливіших з них відносяться механічні та динамічні властивості води та донного ґрунту, температура, світло, розчинені та зважені у воді речовини, активна реакція середовища та деякі інші.
3. Чинники водного середовища. Загальна характеристика. Водне середовище життя, гідросфера, що займає до 71% площі земної кулі, включає близько 1,46 млрд км3 води. Основний обсяг води (приблизно 95%) зосереджений у Світовому океані, левова частка прісних вод – у льодовиках (85%) та підземних водах суші (14%), на озера, водосховища, ставки, болота, річки та струмки доводиться трохи більше 0, 6% від загального обсягу прісної води, що залишаються, 0,35% укладено у ґрунтовій волозі та парах атмосфери.
У водному середовищі мешкають близько 150 000 видів тварин (приблизно близько 7% від загальної кількості на Землі) та 10 000 видів рослин (8%). Отже вода як середовище життя не відрізняється видовою різноманітністю.
У морях і океанах тварина і рослинний світнайбільш різноманітний і багатий в екваторіальній та тропічній зонах. З віддаленням від цих поясів північ і південь якісний склад організмів поступово бідніє. Переважна більшість організмів Світового океану зосереджена біля берегів, переважно у зоні морських узбереж. Відкриті води, розташовані далеко від берегів, є пустельними областями, практично позбавленими життя.
Частка річок, озер, боліт у порівнянні з морями та океанами у біосфері незначна. Незважаючи на меншу біорізноманіття в прісних водахта їх невеликий обсяг, вони мають визначне значення для величезної кількості тварин та рослин, а також для людини, забезпечуючи їх необхідним запасом прісної води.
Сучасну гідросферу можна як як середовище життя. Її мешканці надають активний вплив на кругообіг речовин у біосфері.
Властивості води. Значення води у життєдіяльності організмів визначається переважно її фізичними властивостями. Серед цих властивостей насамперед виділяються термічні – велика теплоємність, висока прихована теплота плавлення та випаровування, низька теплопровідність, розширення перед замерзанням. Завдяки цим властивостям підтримується відносна сталість температурного режиму океанів, що у свою чергу зменшує амплітуду коливань температури на земній поверхні. Розширення води перед замерзанням та аномальні зміни щільності води в інтервалі від 0 до +4° З (саме за такої температури вода має найбільшу щільність, при подальшому підвищенні або зниженні температури води її щільність зменшується) забезпечують перемішування водних мас і перешкоджають промерзанню водойм. Якби цих аномалій льод, що утворився в холодну пору року, опускався б на дно водойм, перетворюючи водні басейни в поклади льоду, що відтають влітку лише з поверхні, де знаходили б притулок тільки ефемерні водні організми.
Вода є чудовим розчинником. Це властивість води та її виняткова рухливість роблять воду основним чинником обміну речовин, як і неорганічної, і органічної природі. У розчиненому стані органічні та неорганічні речовини надходять до споживачів. Водою транспортуються речовини всередині організмів, із водою виділяються продукти розпаду.
завдяки високому поверхневому натягуводи (по її поверхні здатні бігати водомірки), вона утримується на поверхні живих та неживих об'єктів та піднімається по капілярах. Саме ця властивість води забезпечує харчування наземних рослин.
Практична стисливістьводи дозволяє організмам населяти великі глибини.
Завдяки низці оптичних властивостей насамперед, прозоростіУ воді на значних глибинах може йти фотосинтез.
Механіко-динамічні властивості води та ґрунтувизначають умови фіксації та пересування гідробіонтів у просторі. Вони зумовлюють умови плавання пелагічних організмів та його можливості утримання у тих чи інших просторових ділянках водойми. Бентосні організми в залежності від механіко-динамічних властивостей грунту мають різні можливості закапування в субстрат, фіксації на ньому та пересування. Недостатня стабільність ґрунту – перешкода для існування бентосних організмів (неможливість будівництва притулків, механічні пошкодження, вимивання в товщу води, поховання під товщею опадів, що осідають тощо). Рух води - винос за межі біотопів зі сприятливими умовами проживання та навпаки.
Екологічні групи водних організмів. На основі способу життя серед водних організмів виділяють декілька груп:
– бентос(Від грец. benthos– глибина) – прикріплені до ґрунту, що лежать на ньому або живуть у товщі опадів організми. Представлений фітобентосом, зообентосом та бактеріобентосом. У зообентосі за способом харчування виділяють хижаків, фільтраторів, ґрунтоєдів і т.д.;
– перифітон(Від грец. peri- навколо, біля) - тварини та рослини, або прикріплені, або утримувані за стебла та листя вищих рослин або будь-які поверхні, що височіють над дном і плавають за течією;
– планктон(Від грец. planktos- ширяючий, блукаючий і on- Суще) - пасивно плаваючі рослинні (фітопланктон) або тварини (зоопланктон) організми, що переміщуються головним чином завдяки течіям;
– нектон(Від грец. nektos– плаваючий) – активні плавці з обтічною формою тіла та розвиненими органами руху, не пов'язані безпосередньо з дном (риби, кальмари, ластоногі, кити тощо);
– нейстон(Від грец. neustos– плаваючий) – співтовариство мікроорганізмів, рослин та тварин дрібних і середніх розмірів, що мешкають біля поверхні води на межі водного та повітряного середовища (над або під поверхневою плівкою, до 5 см у глиб води). Це бактерії, найпростіші, клопи-водомірки, водорості, личинки деяких організмів;
– плейстон(Від грец. pleystikos– плаваючий) – сукупність гідробіонтів, частина тіла яких у воді, а частина – над її поверхнею. До них відносяться вітрильники, деякі сифонофори та членистоногі, ряска.
Мешканці річок звуться потамобіонтів. Ті з них, що живуть лише на протязі, називаються реофілами. Рух води в річках викликає ерозію її ложа, що йде в глибинному та бічному напрямках. Перенесення та седиментація ґрунту змінюють конфігурацію ложа в горизонтальному та вертикальному напрямках. В результаті бічної ерозії річка меандрує, утворюючи закрути, або меандри. Коли річка спрямовує своє русло, тоді викрутини, що відшнурувалися, перетворюються на стариці. В інших випадках від річки можуть відокремлюватися, не втрачаючи з нею зв'язку, закостя, затониі протоки.
за біологічної класифікації, запропонованої А. Тинеманном та Є. Науманном, розрізняють озера евтрофні, оліготрофні та дистрофні(eu – добре, oligos – мало, dis – недостатньо, trophos – їжа). До евтрофним(Висококормним) відносяться неглибокі рівнинні озера з добре вираженою літароллю і багатою рослинністю. Їхня вода багата на поживні солі, грунти містять багато органічної речовини, і на них розвивається багата донна фауна. Оліготрофні(малокормові) озера зазвичай бувають розташовані на кристалічних породах, мають значну глибину і слаборозвинену літароллю. Донні відкладення бідні на органічні речовини, у воді міститься мало поживних солей, відповідно, життя на дні і в товщі води відрізняється бідністю. Дистрофні(недостатньо кормні) озера є неглибокими заболоченими водоймищами з торф'янистими відкладеннями на дні. Торф'янисті відкладення виключають контакт води з ґрунтом і тому в ній міститься дуже мало мінеральних речовин, солей, необхідних харчування рослин. Планктон та бентос у цих озерах розвинений дуже слабо. Мешканці озер називаються лімнобіонтами. Відповідно до членування бенталі виділяються форми.
Активна реакція середовищає властивістю, що обумовлюється присутністю іонів Н + і ОН - . Як відомо, частина молекул води дисоціює на ці іони. Якщо концентрації іонів Н+ та ОН – рівні, вода буде нейтральною. Зі збільшенням вмісту іонів Н+ та ОН – вода буде відповідно кислою (Н+) або лужною (ОН-). Зазвичай як показник активної реакції береться не концентрація Н + , та її десятковий логарифм зі зворотним знаком. Ця величина, яка називається водневим показником, позначається символом рН. Якщо рН менше 7, вода кисла, більше 7 - лужна, для нейтральної води рН дорівнює 7. Ступінь дисоціації води залежить від температури: зі зниженням останньої вміст Н + падає, а з підвищенням відповідно збільшується. Отже, висновок про активну реакцію води треба робити з урахуванням температури.
Активна реакція природних вод є досить стійкою. Проте в одному і тому ж водоймі рН протягом доби може коливатися на 2 одиниці і більше: вночі знижуватися в результаті підкислення води вуглекислим газом, що виділяється в процесі дихання, вдень підвищується за рахунок споживання ЗІ фотосинтезуючими рослинами. У ґрунтах озер і боліт рН зазвичай дещо нижче 7, в океанічних опадах він часто буває зрушений у лужний бік. Залежно від відношення до різних концентрацій водневих та гідроксильних іонів гідробіонти поділяються на стеноіонних, що мешкають у водах з коливанням рН 5–6 одиниць, та еврійоннихвитримують великі зміни цього фактора.
Концентрація водневих іонів як визначає межі поширення гідробіонтів, а й впливає характер їх життєдіяльності.
Окислювально-відновний потенціал характеризує собою умови протікання серед окислювальних і відновлювальних процесів, яких залежить багато сторін життєдіяльності гидробионтов особливо мікроорганізмів. Нагадаю, що окислювально-відновна реакція полягає в тому, що одна речовина, віддаючи свої електрони та заряджаючи позитивно, окислюється, а інша, набуваючи електрони та заряджаючи негативно, відновлюється. В результаті між ними виникає різницю електричних потенціалів. Чим більша ця різниця, тим вища окисна здатність середовища. Вона визначається концентрацією молекулярного водню у воді, а також станом таких рівноважних систем, як Fe(3) – Fe(2), Mn(4) – Mn(2), S(2) – S(1), а також іншими елементами із змінною валентністю.
Таким чином, умови життя у воді своєрідні. Велике впливом геть розподіл організмів надають світло, температура, течії, тиску, активна реакція середовища, розчинені гази (насамперед кисень) і солі. Морські та континентальні води різко відрізняються за умовами життя. Морська вода є більш сприятливим середовищем, близьким до фізіологічно врівноваженого розчину. Тому її мешканці осмотично відкриті. Континентальні води мають несприятливі умови життя, їх мешканці осмотично закриті.
Адаптивні особливості водних рослин . Водні рослини на відміну від наземних поглинають вологу та мінеральні солі безпосередньо з навколишньої води, тому їхня організація має свої особливості. У них слабо розвинені провідні тканини, а також коренева система. Оскільки коріння служать в основному для прикріплення до підводного субстрату, вони позбавлені кореневих волосків. Потужний розвиток кореневої системи у деяких з них – латаття, кубичок – забезпечує вегетативне розмноження та запасання деяких речовин.
Головною структурною особливістю гідрофітів є наявність великих міжклітин і порожнин, що створюють особливу повітряну тканину, яка забезпечує плавучість органів. Підводні гідрофіти відрізняються від надводних відсутністю функціонуючих продихів, тонким розсіченим листям, слабким розвитком механічних тканин. Інтенсивний газообмін при нестачі у воді розчиненого кисню забезпечується дуже довгими і тонкими стеблами і листям, покриви яких легко проникні для кисню, або сильною розчленованістю листя.
У ряду рослин розвинена гетерофілія (різнолистість). У латаття і кубиків плаваюче листя сильно відрізняється від занурених - їх верхня поверхня щільна і шкіряста, з великою кількістю продихів, що сприяє кращому газообміну з повітрям, на нижній стороні продихів немає.
Через низьку температуру води, що негативно впливає на органи розмноження, і високої щільності середовища, що ускладнює перенесення пилку, занурені у воду рослини розмножуються вегетативним шляхом. Однак багато хто з них виносить квітконосні стебла в повітряне середовище і розмножується статевим шляхом. Їх пилок, плоди та насіння розносяться вітром та поверхневими течіями. Поверхневі течії використовують і прибережні рослини. Їхні плоди мають високу плавучість і можуть тривалий час, перебуваючи у воді, не втрачати схожості.
Адаптивні особливості водних тварин. Адаптації тварин до водного середовища більш різноманітні, ніж рослини. Для тварин, що мешкають у товщі води, характерні, перш за все, пристосування, що збільшують їх плавучість і дозволяють їм протистояти руху води та течії. У дрібних форм спостерігається редукція скелетних утворень. Вони мають пористі раковини або порожнисті всередині голки скелетів. Питома густина тіла зменшується за рахунок наявності води, повітря або жиру в тканинах.
Для тварин, що пасивно плавають у товщі води, характерно також збільшення питомої поверхні тіла. Це досягається ущільненням тіла, утворенням усіляких шипів, виростів.
Активне плавання здійснюється за допомогою вій, джгутиків, а також згинання тіла. Набуло поширення плавання реактивним способом за рахунок енергії струменя води, що викидається. Так, деякі кальмари розвивають швидкість 40–50 км/год. У великих тварин є спеціалізовані кінцівки – плавці, ласти. Тіло у таких тварин покрите слизом і має форму, що обтікає.
Прісноводні тварини при пересуванні використовують поверхневу плівку води. Нею вільно бігають жуки-вертячки, клопи-водомірки. Їхні покриви не змочуються водою, а кінцівки мають особливу будову.
Донні організми, навпаки, виробляють пристосування, що зменшують плавучість і дозволяють утримуватись на дні навіть у швидкоплинних водах. Добре відомі важкі раковини молюска тридакни, які вільно лежать на малій глибині і утримуються на рифах завдяки своїй масі.
Тільки водному середовищі зустрічаються тварини, які ведуть нерухомий спосіб життя. З прикріплених до ґрунту найбільш відомі губки, гідроїдні та коралові поліпи, морські лілії, двостулкові молюски та ін.
Для водяних тварин має значення тиск середовища. Серед еврибатних – що мешкають і при високому і при низькому тиску – виділяються голотурії, що живуть на глибині від 100 до 9000 м. Серед стенобатних – морські лілії, погонофори, що мешкають на глибинах від 3000 до 10 000 м. Для глибоководних тварин характерний слабкий вапняного скелета, редукція органів зору, посилення розвитку дотикових рецепторів, відсутність пігментації тіла або навпаки темне забарвлення.
4.1. Водне середовище проживання. Специфіка адаптації гідробіонтів
Вода як довкілля має ряд специфічних властивостей, таких, як велика щільність, сильні перепади тиску, відносно малий вміст кисню, сильне поглинання сонячних променів та ін. Водойми та окремі їх ділянки різняться, крім того, сольовим режимом, швидкістю горизонтальних переміщень , змістом завислих частинок. Для життя придонних організмів мають значення властивості ґрунту, режим розкладання органічних залишків тощо. Тому разом з адаптаціями до загальних властивостей водного середовища її мешканці повинні бути пристосовані і до різноманітних приватних умов. Мешканці водного середовища отримали в екології загальну назву гідробіонтів. Вони населяють Світовий океан, континентальні водойми і підземні води. У будь-якій водоймі можна виділити різні за умовами зони.
4.1.1. Екологічні зони Світового океану
В океані і морях, що входять до нього, розрізняють насамперед дві екологічні області: товщу води –пелагіаль та дно –бенталь (рис. 38). Залежно від глибини бенталь ділиться насубліторальну зону - область плавного зниження суші до глибини приблизно 200 м, батіальну - область крутого схилу і абісальну зону– область океанічного ложа із середньою глибиною 3–6 км. Ще глибші області бенталі, що відповідають западинам океанічного ложа, називають ультраабісаллю.Кромка берега, що заливається під час припливів, називається літораллю. Вище рівня припливів частина берега, що зволожується бризками прибою, отримала назву супраліторалі.
Мал. 38. Екологічні зони Світового океану Природно, що, наприклад, жителі субліторалі живуть за умов щодо
невисокого тиску, денного сонячного освітлення, часто досить значних змінтемпературний режим. Мешканці абісальних та ультраабісальних глибин існують у темряві, при постійній температурі та жахливому тиску в кілька сотень, а іноді й близько тисячі атмосфер. Тому одна лише вказівка на те, в якій зоні бенталі мешкає той чи інший вид організмів, вже говорить про те, якими загальними екологічними властивостями він повинен мати. Все населення дна океану отримало назву бентоса.
Н. М. Чернова, А. М. Билова. «Загальна екологія»
Організми, що мешкають у товщі води, або пелагіалі, відносяться до пелагосу. Пелагіаль також ділять на вертикальні зони, що відповідають по глибині зонам бенталі: епіпе-
лагіаль, батипелагіаль, абісопелагіаль.Нижня межа епіпелагіалі (не більше 200 м)
визначається проникненням сонячного світла у кількості, достатній для фотосинтезу. Фотосинтезуючі рослини глибше цих зон не можуть існувати. У сутінкових батіальних і повних мороку абісальних глибинах мешкають лише мікроорганізми та тварини. Різні екологічні зони виділяються і в усіх інших типах водойм: озерах, болотах, ставках, річках і т. д. Різноманітність гідробіонтів, які освоїли всі ці місця проживання, дуже велика.
4.1.2. Основні властивості водного середовища
Щільність води - це фактор, що визначає умови пересування водних органів.
нізмів і тиск на різних глибинах. Для дистильованої води густина дорівнює 1 г/см3 при 4 °C. Щільність природних вод, що містять розчинені солі, може бути більшою,
до 1,35 г/см3. Тиск зростає з глибиною приблизно в середньому на 1 · 105 Па (1 атм) на кожні 10 м-коду.
У зв'язку з різким градієнтом тиску у водоймах гідробіонти загалом значно еврібатніші порівняно із сухопутними організмами. Деякі види, поширені різних глибинах, переносять тиск від кількох до сотень атмосфер. Наприклад, голотурії роду Elpidia, черв'яки Priapulus caudatus мешкають від прибережної зонидо ультраабіссалі. Навіть прісноводні жителі, наприклад інфузорії-туфельки, сувойки, жуки-
плавунці та ін, витримують у досвіді до 6 · 107 Па (600 атм).
Однак багато жителів морів і океанів щодо стенобатних і приурочені до певних глибин. Стенобатність найчастіше властива мілководним і глибоководним видам. Тільки на літоралі мешкають кільчастий хробак песшкіріл Arenicola, молюски морські блюдечки (Patella). Багато риб, наприклад з групи вудильників, головоногі молюски, ракоподібні, погонофори, морські зірки та ін зустрічаються лише на великих
глибин при тиску не менше 4 · 107 - 5 · 107 Па (400-500 атм).
Щільність води забезпечує можливість спиратися на неї, що особливо важливо для бесскелетних форм. Щільність середовища служить умовою ширяння в воді, і багато гідробіонти пристосовані саме до цього способу життя. Зважені, парячі у воді організми об'єднують у особливу екологічну групу гідробіонтів – планктон (планктос)
– ширяючий).
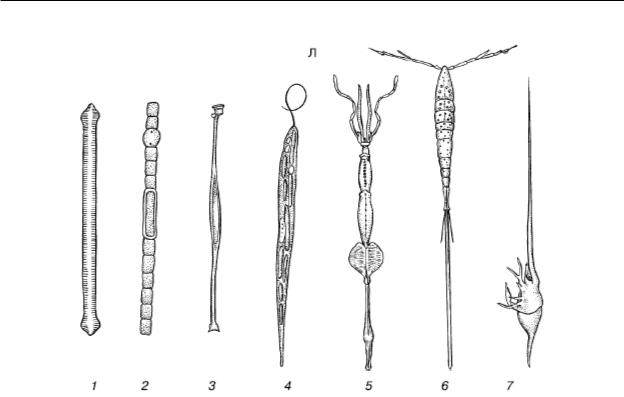
Н. М. Чернова, А. М. Билова. «Загальна екологія»
Мал. 39. Збільшення відносної поверхні тіла у планктонних організмів (за С. А. Зерновим, 1949):
A – паличкоподібні форми:
1 – діатомея Synedra;
2 – ціанобактерія Aphanizomenon;
3 – перидінева водорість Amphisolenia;
4 – Euglena acus;
5 – головоногий молюск Doratopsis vermicularis;
6 – веслоногий рачок Setella;
7 – личинка Porcellana (Decapoda)
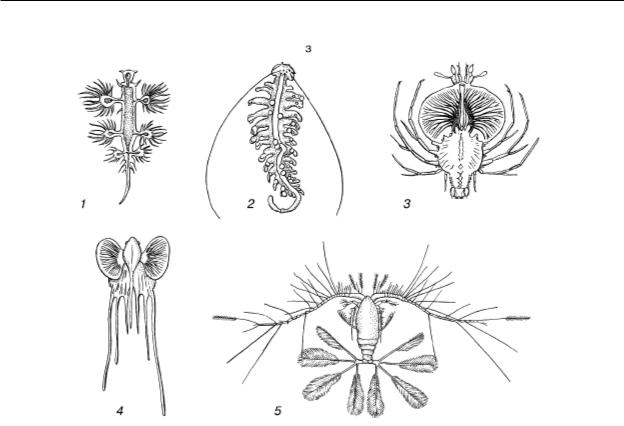
Н. М. Чернова, А. М. Билова. «Загальна екологія»
Б - розчленовані форми:
1 – молюск Glaucus atlanticus;
2 – хробак Tomopetris euchaeta;
3 – личинка раку Palinurus;
4 - личинка риби морського чорта Lophius;
5 – веслоногий рачок Calocalanus pavo
У складі планктону – одноклітинні та колоніальні водорості, найпростіші, медузи, сифонофори, гребневики, крилоногі та кіленогі молюски, різноманітні дрібні рачки, личинки донних тварин, ікра та мальки риб та багато інших (рис. 39). Планктонні організми мають багато подібних адаптацій, що підвищують їх плавучість і перешкоджають осіданню на дно. До таких пристроїв відносяться: 1) загальне збільшення відносної поверхні тіла за рахунок зменшення розмірів, сплющеності, подовження, розвитку численних виростів або щетинок, що збільшує тертя про воду; 2) зменшення щільності за рахунок редукції скелета, накопичення в тілі жирів, бульбашок газу тощо. У діатомових водоростей запасні речовини відкладаються не у вигляді важкого крохмалю, а у вигляді жирових крапель. Ночосвітка Noctiluca відрізняється такою великою кількістю газових вакуолей і крапельок жиру в клітині, що цитоплазма в ній має вигляд тяжів, що зливаються тільки навколо ядра. Повітряні камери є і в сифонофор, ряду медуз, планктонних черевоногих молюсків та ін.
Водорості (фітопланктон) парять у воді пасивно, більшість планктонних тварин здатне до активного плавання, але в обмежених межах. Планктонні організми що неспроможні долати течії і переносяться ними великі відстані. Багато видів зоопланктону здатні, однак, до вертикальних міграцій у товщі води на десятки і сотні метрів як за рахунок активного пересування, так і за рахунок регулювання плавучості свого тіла. Особливий різновид планктону становить екологічна група нейстону («нейн» – плавати) – мешканці поверхневої плівки води на кордоні з повітряним середовищем.
Н. М. Чернова, А. М. Билова. «Загальна екологія»
Щільність і в'язкість води сильно впливають на можливість активного плавання. Тварин, здатних швидкого плавання і подолання сили течій, об'єднують в екологічну групу нектону («нектос» – плаваючий). Представники нектону – риби, кальмари, дельфіни. Швидкий рух у водній товщі можливо лише при наявності обтічної форми тіла і сильно розвиненою мускулатури. Торпедоподібна форма виробляється у всіх хороших плавців незалежно від їхньої систематичної приналежності та способу руху у воді: реактивного, за рахунок згинання тіла, за допомогою кінцівок.
Кисневий режим.У насиченій киснем воді вміст його не перевищує 10 мл в 1 л, це в 21 разів нижче, ніж в атмосфері. Тому умови дихання гідробіонтів значно ускладнені. Кисень надходить в воду в основному за рахунок фотосинтетичної діяльності водоростей і дифузії з повітря. Тому верхні шари водної товщі, як правило, багатше цим газом, ніж нижні. З підвищенням температури і солоності води концентрація в ній кисню знижується. У шарах, сильно заселених тваринами та бактеріями, може створюватися різкий дефіцит О2 через посилене його споживання. Наприклад-
мір, у Світовому океані багаті життям глибини від 50 до 1000 м характеризуються різким погіршенням аерації – вона у 7-10 разів нижча, ніж у поверхневих водах, населених фітопланктоном. Близько дна водойм умови можуть бути близькі до анаеробних.
Серед водних жителів багато видів, здатних переносити широкі коливання вмісту кисню у воді, аж до майже повної його відсутності (евріоксибіонти - "окси" - кисень, "біонт" - мешканець). До них відносяться, наприклад, прісноводні олігохети Tubifex tubifex, черевоногих молюсків Viviparus viviparus. Серед риб дуже слабке насичення води киснем можуть витримувати сазан, лин, карасі. Разом з тим ряд видів стеноксібіонтни– вони можуть існувати лише за досить високому насиченні води киснем (райдужна форель, кумжа, гольян, війний черв'як Planaria alpina, личинки поденок, веснянок та інших.). Багато видів здатні при нестачі кисню впадати в неактивний стан аноксибіоз і таким чином переживати несприятливий період.
Дихання гідробіонтів здійснюється через поверхню тіла, або через спеціалізовані органи - зябра, легені, трахеї. При цьому покриви можуть бути додатковим органом дихання. Наприклад, риба в'юн через шкіру споживає в середньому до 63% кисню. Якщо через покриви тіла відбувається газообмін, вони дуже тонкі. Дихання полегшується збільшенням поверхні. Це досягається в ході еволюції видів освітою різних виростів, ущільненням, подовженням, загальним зменшенням розмірів тіла. Деякі види за нестачі кисню активно змінюють величину дихальної поверхні. Хробаки Tubifex tubifex сильно витягають тіло в довжину; гідри та актинії – щупальці; голкошкірі – амбулакральні ніжки. Багато сидячих і малорухливих тварин оновлюють навколо себе воду, або створюючи її спрямований струм, або коливальними рухами сприяючи її перемішування. Двостулковим молюскам для цієї мети служать вії, що вистилають стінки мантійної порожнини; ракоподібним – робота черевних чи грудних ніжок. П'явки, личинки комарів-дзвінців (мотиль), багато олігохетів колишають тіло, висунувшись із ґрунту.
У деяких видів зустрічається комбінування водного та повітряного дихання. Такі двоякодишачі риби, сифонофори дискофанти, багато легеневі молюски, ракоподібні Gammarus lacustris та ін. Вторинні тварини зберігають зазвичай атмосферний тип дихання як більш вигідний енергетично і потребують тому в контактах з повітряним середовищем, наприклад ластоногие, китоподібні, вод.
Нестача кисню у воді призводить іноді до катастрофічних явищ - заморів, що супроводжуються загибеллю безлічі гідробіонтів.
Н. М. Чернова, А. М. Билова. «Загальна екологія»
ються утворенням на поверхні водойм льоду та припиненням контакту з повітрям; літні – підвищенням температури води та зменшенням внаслідок цього розчинності кисню.
Часта загибель риб та багатьох безхребетних взимку характерна, наприклад, для нижньої частини басейну річки Обі, води якої, що стікають із заболочених просторів ЗахідноСибірської низовини, вкрай бідні на розчинений кисень. Іноді замори виникають і морях.
Крім нестачі кисню, замори можуть бути викликані підвищенням концентрації у воді токсичних газів – метану, сірководню, СО2 та ін., що утворюються в результаті роз-
кладення органічних матеріалів на дні водойм.
Сольовий режим. Підтримка водного балансу гідробіонтів має свою специфіку. Якщо для наземних тварин і рослин найбільш важливе забезпечення організму водою в умовах її дефіциту, то для гідробіонтів не менш істотно підтримка певної кількості води в тілі при її надлишку навколишньому середовищі. Зайва кількість води в клітинах призводить до зміни в них осмотичного тиску і порушення найважливіших життєвих функцій.
Більшість водних мешканців пойкілосмотічни:осмотичний тиск в їх тілі залежить від солоності навколишнього води. Тому для гідробіонтів основний спосіб підтримувати свій сольовий баланс - це уникати місць існування з невідповідною солоністю. Прісноводні форми не можуть існувати в морях, морські - не переносять опріснення. Якщо солоність води піддається змін, тварини переміщаються у пошуках сприятливого середовища. Наприклад, при опрісненні поверхневих шарів моря після сильних дощів радіолярії, морські рачки Calanus та інші спускаються на глибину до 100 м. гомойосмотіческімвидам, зберігаючи постійне осмотичний тиск в тілі незалежно від концентрації солей у воді.
У прісноводних видів соки тіла гіпертонічні по відношенню до навколишньої води. Їм загрожує зайве обводнення, якщо не перешкоджати надходженню або не видаляти надлишок води з тіла. У найпростіших це досягається роботою виділень вакуолей, у багатоклітинних - видаленням води через систему виділення. Деякі інфузорії кожні 2-2,5 хв виділяють кількість води, що дорівнює об'єму тіла. На відкачування надлишкової води клітина витрачає дуже багато енергії. З підвищенням солоності робота вакуолей уповільнюється. Так, у туфельок Paramecium при солоності води 2,5% вакуоль пульсує з інтервалом в 9 с, при 5% - 18 с, при 7,5% - 25 с. При концентрації солей 17,5% вакуоль перестає працювати, так як різниця осмотичного тиску між клітиною і зовнішнім середовищем зникає.
Якщо вода гіпертонічна стосовно рідин тіла гідробіонтів, їм загрожує зневоднення внаслідок осмотичних втрат. Захист від зневоднення досягається підвищенням концентрації солей також у тілі гідробіонтів. Зневодненню перешкоджають непроникні для води покриви гомойосмотичних організмів - ссавців, риб, вищих раків, водних комах та їх личинок.
Багато пойкілосмотіческіе види переходять до неактивного стану - анабіозу в результаті дефіциту води в тілі при зростанні солоності. Це властиво видам, що мешкають у калюжах морської води та на літоралі: коловраткам, джгутиковим, інфузоріям, деяким рачкам, чорноморським поліхетам Nereis divesicolor та ін. Сольовий анабіоз – засіб переживати несприятливі періоди в умовах змінної солоності води.
Істинно евригалінних видів, здатних в активному стані жити як у прісній, так і в солоній воді, серед водних жителів не так багато. В основному це види, що населяють естуарії річок, лимани та інші солонуваті водойми.
Н. М. Чернова, А. М. Билова. «Загальна екологія»
Температурний режимводойм більш стійкий, ніж на суші. Це пов'язано з фізичними властивостямиводи, насамперед високою питомою теплоємністю, завдяки якій одержання чи віддача значної кількості тепла не викликає надто різких змін температури. Випаровування води з поверхні водойм, при якому витрачається близько 2263,8 Дж/г, перешкоджає перегріванню нижніх шарів, а утворення льоду, при якому виділяється теплота плавлення (333,48 Дж/г), уповільнює їхнє охолодження.
Амплітуда річних коливань температури у верхніх шарах океану трохи більше 10–15 °C, у континентальних водоймах – 30–35 °C. Глибокі шари води відрізняються сталістю температури. У екваторіальних водах середньорічна температура поверхневих шарів +(26-27) ° С, у полярних - близько 0 ° C і нижче. У гарячих наземних джерелах температура води може наближатися до +100 °C, а підводних гейзерах при високому тиску на дні океану зареєстрована температура +380 °C.
Таким чином, в водоймах існує досить значна різноманітність температурних умов. Між верхніми шарами води з вираженими в них сезонними коливаннями температури та нижніми, де тепловий режим постійний, існує зона температурного стрибка, або термокліна. Термоклін різкіше виражений у теплих морях, де сильніший перепад температури зовнішніх та глибинних вод.
У зв'язку з більш стійким температурним режимом води серед гідробіонтів в значно більшій мірі, ніж серед населення суші, поширена стенотермним. Евритермные види зустрічаються переважно у дрібних континентальних водоймах і літоралі морів високих і помірних широт, де значні добові і сезонні коливання температури.
Світловий режим. Світла в воді набагато менше, ніж в повітрі. Частина падаючих на поверхню водойми променів відбивається в повітряне середовище. Відображення тим сильніше, чим нижче положення Сонця, тому день під водою коротше, ніж на суші. Наприклад, літній день біля острова Мадейра на глибині 30 м – 5 год, а на глибині 40 м лише 15 хв. Швидке зменшення кількості світла з глибиною пов'язане з поглинанням його водою. Промені з різною довжиноюхвилі поглинаються неоднаково: червоні зникають вже неподалік поверхні, тоді як синьо-зелені проникають значно глибше. Ті, що згущуються з глибиною сутінки в океані мають спочатку зелений, потім блакитний, синій і синьо-фіолетовий колір, змінюючись нарешті постійним мороком. Відповідно змінюють один одного з глибиною зелені, бурі та червоні водорості, спеціалізовані на уловлюванні світла з різною довжиною хвилі.
Забарвлення тварин змінюється із глибиною так само закономірно. Найбільш яскраво та різноманітно забарвлені жителі літоральної та субліторальної зон. Багато глибинні організми, подібно до печерних, не мають пігментів. У сутінковій зоні широко поширене червоне забарвлення, яке є додатковим до синьо-фіолетового світла на цих глибинах. Додаткові за кольором промені найповніше поглинаються тілом. Це дозволяє тваринам ховатися від ворогів, тому що їх червоний колір у синьо-фіолетових променях візуально сприймається як чорний. Червоне забарвлення й у таких тварин сутінкової зони, як морський окунь, червоний корал, різні ракоподібні та інших.
У деяких видів, що мешкають біля поверхні водойм, очі поділяються на дві частини з різною здатністю до заломлення променів. Одна половина ока бачить у повітрі, інша – у воді. Така «чотирьокість» характерна для жуків-вертячок, американської рибки Anableps tetraphthalmus, одного з тропічних видів морських собачок Dialommus fuscus. Ця рибка при відливах сидить у поглибленнях, виставляючи частину голови із води (див.
Поглинання світла тим сильніше, чим менша прозорість води, яка залежить від кількості зважених у ній частинок.

Н. М. Чернова, А. М. Билова. «Загальна екологія»
Прозорість характеризують граничною глибиною, на якій ще видно білий диск, що спеціально опускається, діаметром близько 20 см (диск Секкі). Найпрозоріші води – у Саргасовому морі: диск видно до глибини 66,5 м. У Тихому океані диск Секкі видно до 59 м, в Індійському – до 50, у дрібних морях – до 5-15 м. Прозорість рік у середньому 1–1 ,5 м, а в найкаламутніших річках, наприклад у середньоазіатських Амудар'ї та Сирдар'ї, всього кілька сантиметрів. Кордон зони фотосинтезу тому сильно варіює у різних водоймах. У найчистіших водах еуфотична зона, або зона фотосинтезу, простягається до глибин не понад 200 м, сутінкова, або дисфотична, зона займає глибини до 1000-1500 м, а глибше, вафотичну зону, сонячне світло не проникає зовсім.
Кількість світла у верхніх шарах водойм сильно змінюється в залежності від широти місцевості і від пори року. Довгі полярні ночі сильно обмежують час, придатний для фотосинтезу, в арктичних та приантарктичних басейнах, а льодовий покрив ускладнює доступ світла взимку у всі водойми, що замерзають.
У темних глибинах океану в якості джерела зорової інформації організми використовують світло, що випускається живими істотами. Світіння живого організму отримало назву біолюмінесценції.Види, що світяться, є майже у всіх класах водних тварин від найпростіших до риб, а також серед бактерій, нижчих рослин і грибів. Біолюмінесценція, мабуть, багаторазово виникала у різних групах на різних етапах еволюції.
Хімію біолюмінесценції зараз досить добре вивчено. Реакції, що використовуються для генерації світла, різноманітні. Але у всіх випадках це окиснення складних органічних сполук (люциферинів) за допомогою білкових каталізаторів (люцифераз). Люциферини та люциферази у різних організмів мають неоднакову структуру. У результаті реакції надлишкова енергія збудженої молекули люциферину виділяється як квантів світла. Живі організми випромінюють світло імпульсами, зазвичай у відповідь роздратування, що надходять із зовнішнього середовища.
Світіння може і не відігравати особливої екологічної ролі в житті виду, а бути побічним результатом життєдіяльності клітин, як, наприклад, бактерій або нижчих рослин. Екологічну значущість воно отримує лише у тварин, які мають досить розвинену нервову систему та органи зору. У багатьох видів органи світіння набувають дуже складної будови із системою відбивачів і лінз, що підсилюють випромінювання (рис. 40). Ряд риб і головоногих молюсків, нездатних генерувати світло, використовують симбіотичних бактерій, які розмножуються спеціальних органах цих тварин.